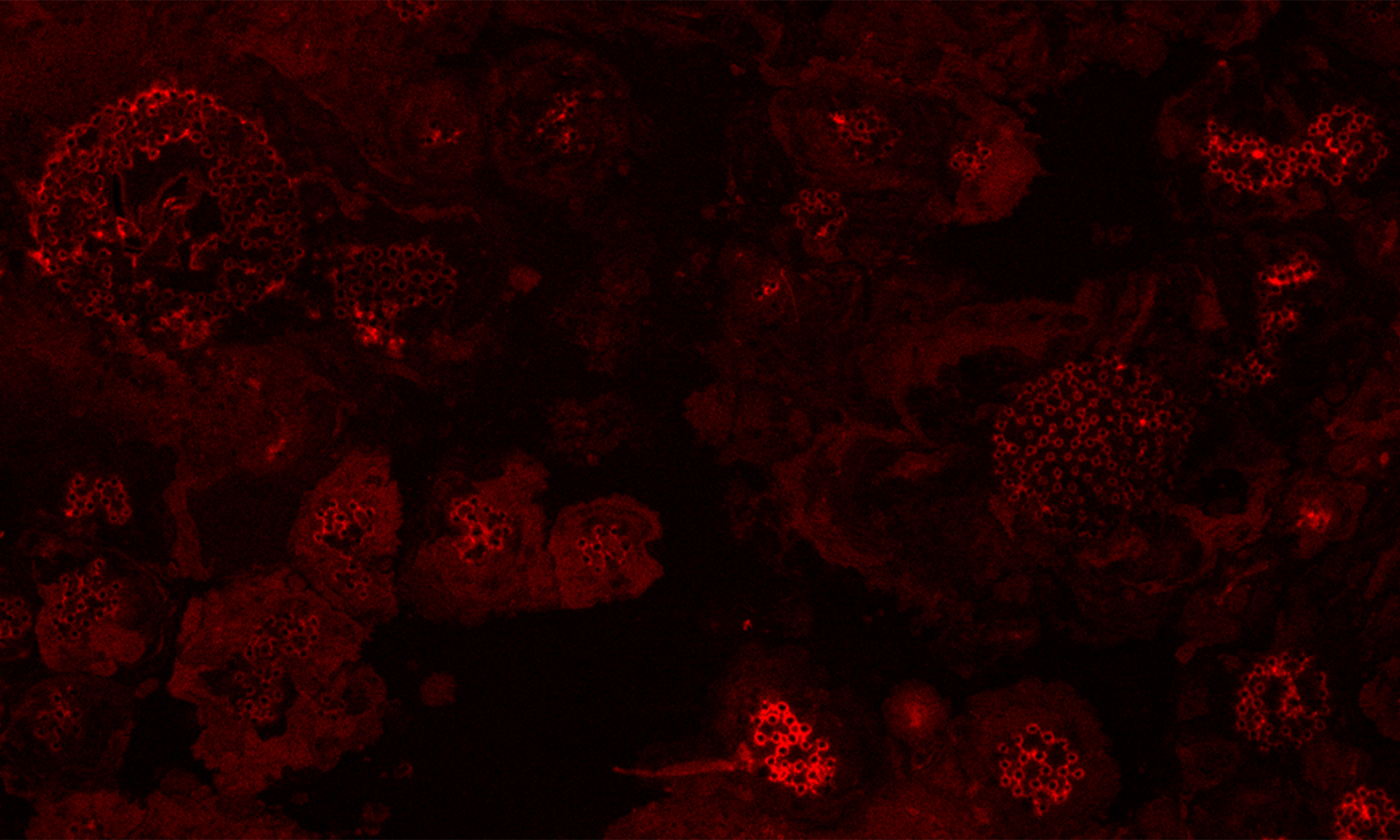
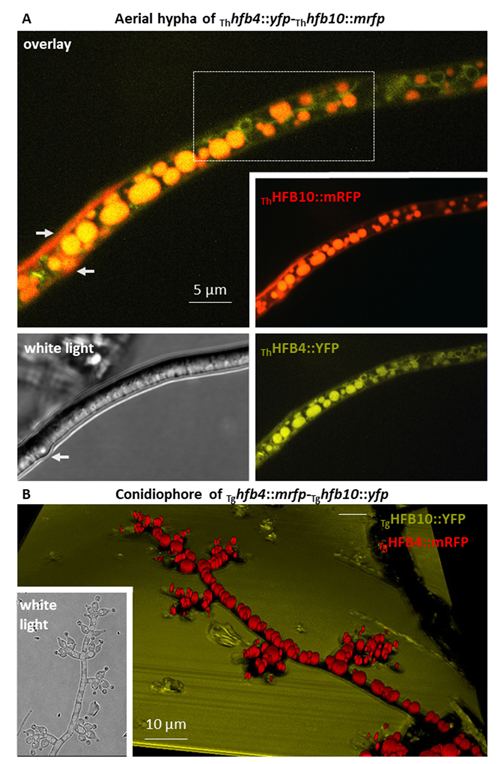
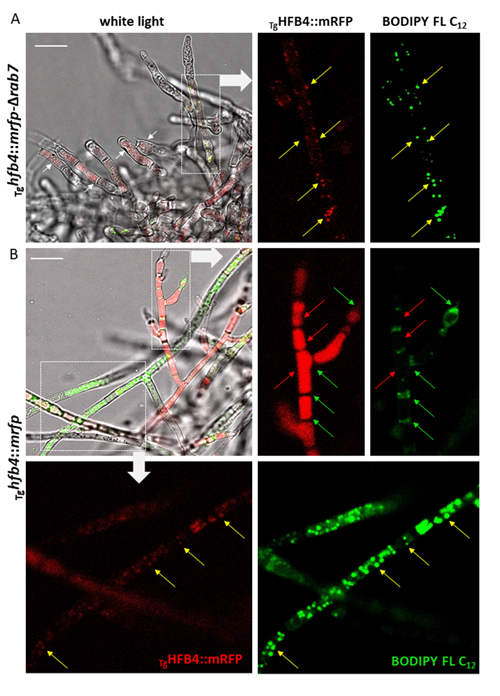
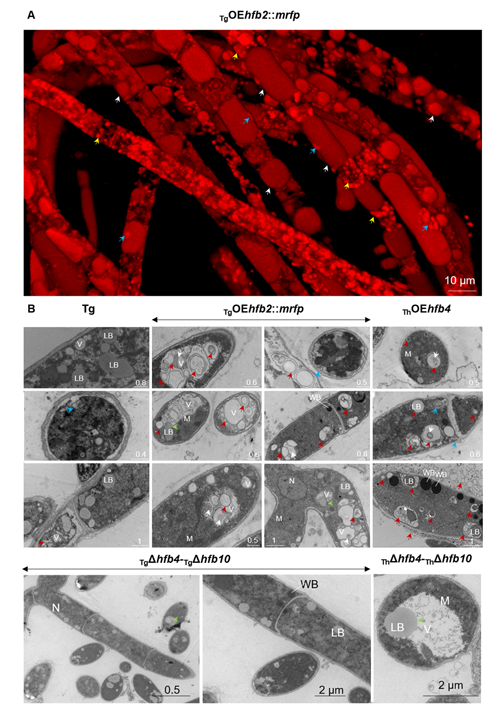
Animated 3D reconstructions of extracellular HFB-enriched matrices coating sporulating Trichoderma colonies.
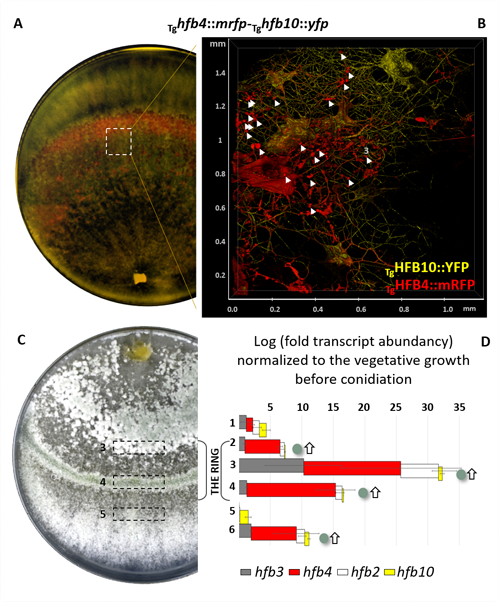
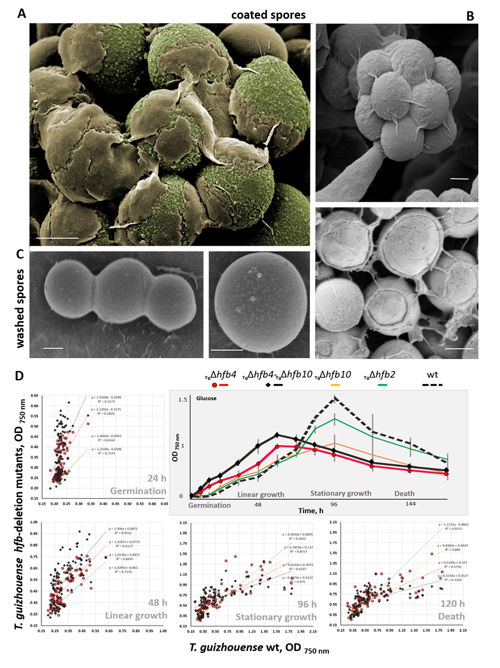
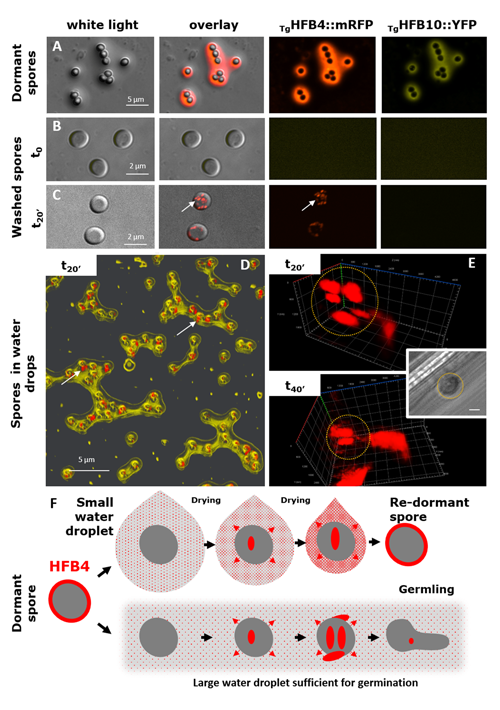
Higher fungi can rapidly produce large numbers of spores suitable for aerial dispersal. The efficiency of the dispersal and spore resilience to abiotic stresses correlate with their hydrophobicity provided by the unique amphiphilic and superior surface-active proteins – hydrophobins (HFBs) – that self-assemble at hydrophobic/hydrophilic interfaces and thus modulate surface properties. Using the HFB-enriched mold Trichoderma (Hypocreales, Ascomycota) and the HFB-free yeast Pichia pastoris (Saccharomycetales, Ascomycota), we revealed that the rapid release of HFBs by aerial hyphae shortly prior to conidiation is associated with their intracellular accumulation in vacuoles and/or lipid-enriched organelles. The occasional internalization of the latter organelles in vacuoles can provide the hydrophobic/hydrophilic interface for the assembly of HFB layers and thus result in the formation of HFB-enriched vesicles and vacuolar multicisternal structures (VMSs) putatively lined up by HFBs. These HFB-enriched vesicles and VMSs can become fused in large tonoplast-like organelles or move to the periplasm for secretion. The tonoplast-like structures can contribute to the maintenance of turgor pressure in aerial hyphae supporting the erection of sporogenic structures (e.g., conidiophores) and provide intracellular force to squeeze out HFB-enriched vesicles and VMSs from the periplasm through the cell wall. We also show that the secretion of HFBs occurs prior to the conidiation and reveal that the even spore coating of HFBs deposited in the extracellular matrix requires microscopic water droplets that can be either guttated by the hyphae or obtained from the environment. Furthermore, we demonstrate that at least one HFB, HFB4 in T. guizhouense, is produced and secreted by wetted spores. We show that this protein possibly controls spore dormancy and contributes to the water sensing mechanism required for the detection of germination conditions. Thus, intracellular HFBs have a range of pleiotropic functions in aerial hyphae and spores and are essential for fungal development and fitness.
MycoCosm, KEGG, and other web resources on fungal genetics
Recent literature on fungal genetics (incl. fungal diversity and applications)
Research community
Terminology
Evolution, diversity, and speciation
How will it work?
IrinaDruzhinina WeChat
Register by sending a personal message to FungiG or contact “IrinaDruzhinina” on WeChat. Below please find the QR code.
Get the first task (1/10 of the entire exercise).
Submit your results and questions (Email or Wechat)
Get feedback and answer to your questions.
Repeat until the course is completed.
What is the content of the course?
The WS2021 course will be based on the dynamic list of Trichoderma genes and Trichoderma-associated genes (mainly plant genes studied along with Trichoderma). The next courses may be based on other model fungi.
The tasks for this course are divided into small sets. Each time, a student will get his or her own random set of 2–3 fungal genes. The task will be to search for the genome IDs, protein IDs, function(s), evolutionary history, mutant(s), phenotype(s), GO term(s), KEGG group(s), genomic/chromosome location(s), cluster organization(s), functionally associated gene(s), published gene name(s), host genome(s), orthologue(s), paralogue(s), other homologue(s), patent(s), applied value(s), product(s), and reference(s).
The student is expected to systemically collect associated counts, such as the total number of publications, number of patents, number of Trichoderma spp. studied, etc. Some genes are intensively studied (e.g., cbh1, lae1), and the task will take more effort compared to the others that have only been published once.
What should I know before the course?
Basic eukaryotic microbiology and basic mycology
Basic biochemistry and cell biology
English reading skills
Advanced skills in retrieval of scientific literature (FungiG will provide help)
What is the main challenge of the course?
The concept of a fungal gene, gene definition, gene nomenclature in fungi, inconsistency in research approaches, the diversity of genes, the unequal quality of genome annotations, fungal diversity, and taxonomy.
What exactly should I do to complete the course?
A student is expected to deliver a table (as will be specified in the task) describing the functions and properties of several Trichoderma genes or genes associated with Trichoderma research.
The minimum set of genes is 25 (10 sets of 2–3 genes); the upper limit is 300*.
The advanced version of the course includes the joint (online/offline) seminar with students’ presentations and discussions. The aim of the seminar is to appoint the top ten most studied, most useful, and most controversial genes in Trichoderma, respectively.
* The total number of Trichoderma genes is ~12 000, but the number of genes studied for their function(s) is still meager.
Can my tasks be redundant to the tasks of my colleagues?
Sometimes, yes. The majority of the tasks will be unique. However, the genes used for the tasks that failed or were superficially performed remain in the pool of genes for the course.
Can I add genes to the list?
Yes. Students are welcome to do so. These can also be genes from other fungi that are not yet studied in Trichoderma. Please send your proposals to ISD.
What is the course language?
English.
What is the schedule of the course?
The schedule of the course is flexible. The results can be sent at any time. The feedback will be returned within 72 hours, or the exact time will be specified.
How long will it take? How deep can I go?
The course is designed such that an advanced Ph.D. student working on fungal genetics is expected to spend one day per week for 10 weeks or make it in a block (2 weeks, full time). The minimal workload corresponds to 80–90 working hours or 3 ECTS (European Credit Transfer and Accumulation System, Bologna Process).
As you progress, it should become faster. After you get in shape, you may either spend less time per week or learn more genes.
Can I do the entire course remotely?
Yes.
Who can attend?
The course will present new material to all FungiG members, ranging from master students to Ph.D. candidates, postdocs, and alumni professors working at Nanjing Agricultural University, Shanghai Jiao Tong University, Sun Yat-Sen University, Jiangsu Academy of Agricultural Sciences, and other universities. Students from the TU Wien master program “Biotechnology and Bioanalytics,” are welcome.
Students from the universities or academic institutions that are not listed above, please contact Irina Druzhinina.
Is the course free?
The WS2021 is free of charge but the number of places is limited.
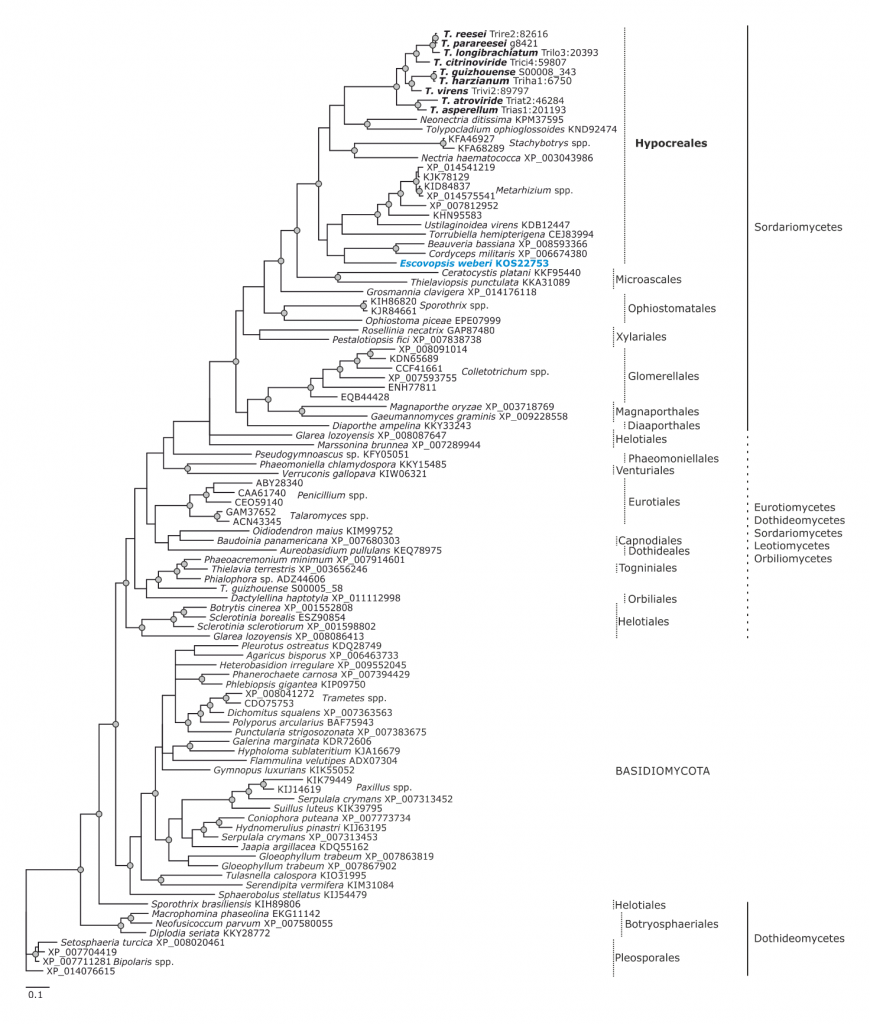
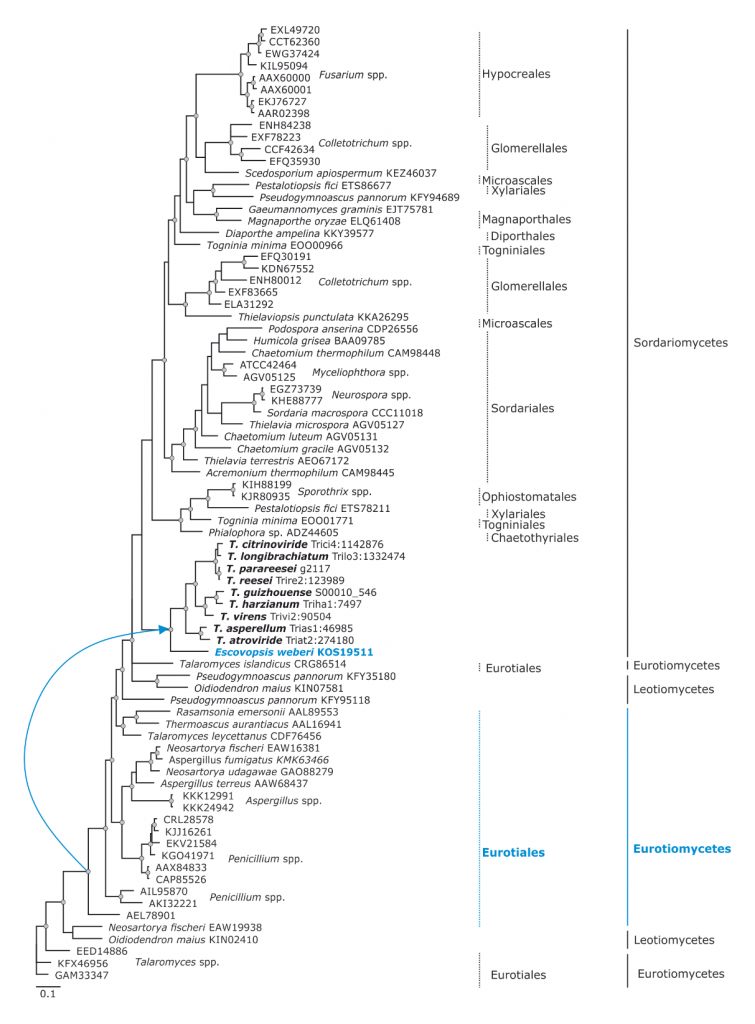
Phylogram based on Dayhoff amino acid substitution model using an alignment containing 70686 characters. Bayesian analysis was run for 1 million mcmc generations and a strict consensus tree was obtained by summarizing 7500 trees, after burning first 25% of obtained 10,000 trees. Mean tree length and variance are 2.425237E+01 and 7.0550690E-02, respectively. Posterior probabilities more than 95% are marked with circular nodes. Dashed vertical bars and non-dashed vertical bars represents the taxonomic order and taxonomic class in the phylum Ascomycota, respectively
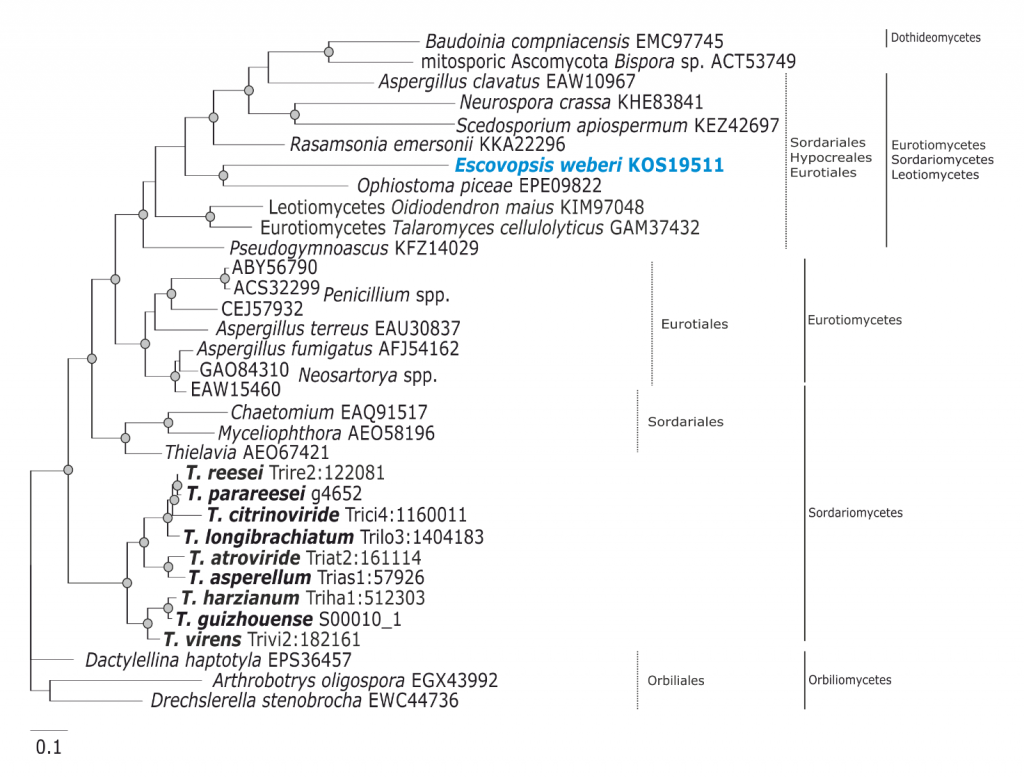
Phylogram based on Dayhoff amino acid substitution model using an alignment containing 38720 characters. Bayesian analysis was run for 1 million mcmc generations and a strict consensus tree was obtained by summarizing 7500 trees, after burning first 25% of obtained 10,000 trees. Mean tree length and variance are 9.480271E+00 and 2.2693320E-02, respectively. Posterior probabilities more than 95% are marked with circular nodes. Dashed vertical bars and non-dashed vertical bars represents the taxonomic order and taxonomic class in the phylum Ascomycota, respectively. LGT inferred by phylogeny and NOTUNG is shown with a blue arrow since it is occuring before Trichoderma and Escovopsis diverged and the respective donors are marked in blue.
Phylogram based on Dayhoff amino acid substitution model using an alignment containing 18546 characters. Bayesian analysis was run for 1 million mcmc generations and a strict consensus tree was obtained by summarizing 7500 trees, after burning first 25% of obtained 10,000 trees. Mean tree length and variance are 7.457580E+00 and 2.1536720E-02, respectively. Posterior probabilities more than 95% are marked with circular nodes. Dashed vertical bars and non-dashed vertical bars represents the taxonomic order and taxonomic class in the phylum Ascomycota, respectively
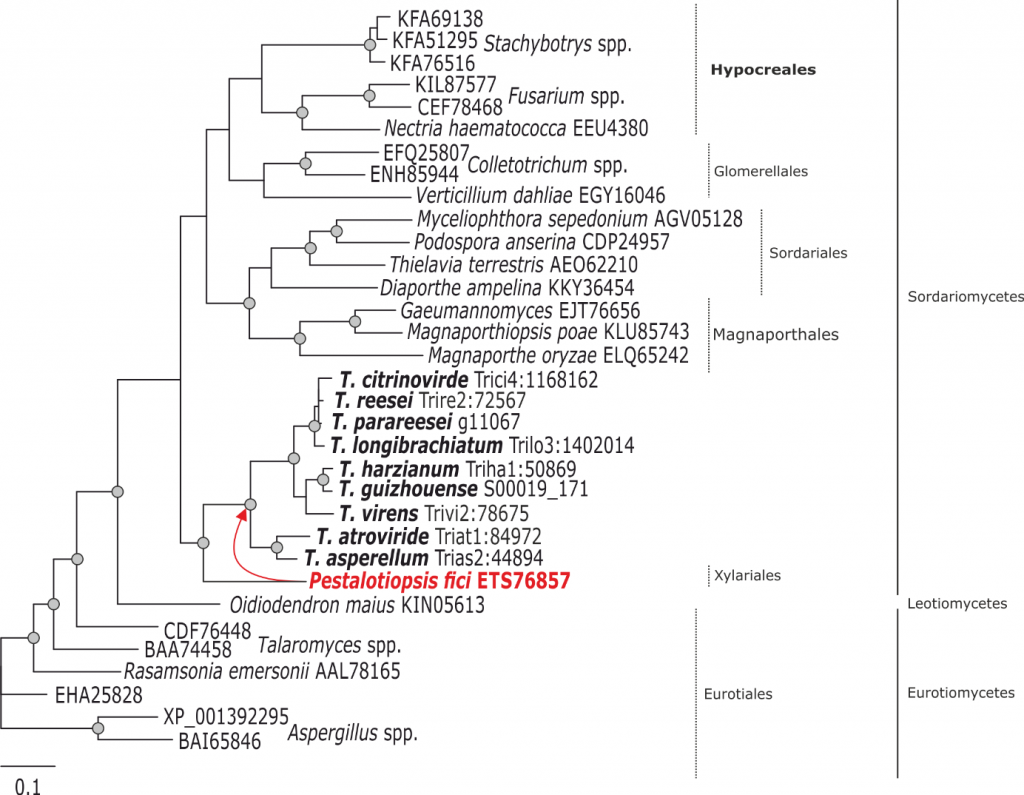
Phylogram based on Dayhoff amino acid substitution model using an alignment containing 20707 characters. Bayesian analysis was run for 1 million mcmc generations and a strict consensus tree was obtained by summarizing 7500 trees, after burning first 25% of obtained 10,000 trees. Mean tree length and variance are 7.269450E+00 and 3.9080650E-02, respectively. Posterior probabilities more than 95% are marked with circular nodes. Dashed vertical bars and non-dashed vertical bars represents the taxonomic order and taxonomic class in the phylum Ascomycota, respectively. LGT inferred by NOTUNG and Phylogeny is shown with a red arrow to the node and the respective donor is marked in red.